 bone collagen pseudomorphs
bone collagen pseudomorphs  the archaeological science laboratory in SCA
the archaeological science laboratory in SCA
Principles of Stable
IsotopeAnalysis
Analytical Methods
Recent Applications
References
Laboratory Staff and
Students
Carbon and nitrogen isotope ratios in human bone may be used to reconstruct prehistoric diet because of differential fractionation, between certain plant groups, of atmospheric carbon dioxide during photosynthesis and of nitrogen during fixation or absorption. There are two stable isotopes each of carbon (12C, 13C) and nitrogen (14N, 15N), with 12C and 14N by far the most common in nature. Small differences in the ratios of these isotopes (12C/13C, 15N /14N) can be measured by isotope ratio mass spectrometry using samples smaller than 1 milligram. High precision isotope measurements are reported using the delta notation (d13C, d15N) relative to internationally recognized standards and are expressed in parts per thousand or per mil (‰).
Experimental data have indicated that different bone tissues reflect different components of the diet (Ambrose and Norr 1993; Tieszen and Fagre 1993). In general, bone collagen is disproportionately produced from the protein portion of the diet, while bone carbonate and tooth enamel carbonate (both a calcium hydroxyphosphate, called apatite) are produced from a mixture of dietary protein, carbohydrates and fats. Stable isotope analysis of both bone collagen and apatite thus permits quantitative estimates of several dietary components. Both bone collagen and bone apatite are constantly being resorbed and replenished, so that their isotopic composition reflects dietary averages over at least the last several years of an individual's life, while the composition of tooth enamel will reflect diet during the age of crown formation.
Since teeth form at different ages (beginning in utero for deciduous teeth, and from 0 to 12 yrs of age for permanent teeth), the analysis of multiple teeth from the same individual may reveal dietary shifts caused by weaning (first the introduction of solid foods and later the cessation of breastfeeding) (Wright & Schwarcz 1998), while significant differences between tooth (juvenile diet) and bone (adult diet) values may be the result of subsistence changes due to migration. Analysis of hair segments also may reveal short-term or seasonal dietary changes (O'Connell & Hedges 1999). Overall, stable isotope analysis of multiple tissues can provide a quantifiable dietary life history of an individual (Sealy et al. 1995), whereas subsistence data from faunal and archaeobotanical remains typically derive from at least multiple individuals, often over multiple generations. While collagen is rarely preserved in bones predating the Upper Paleolithic, and is often badly degraded in hot and moist environments (e.g. lowland Mesoamerica), bone apatite has provided reliable results for the Holocene, and tooth enamel for early hominids (e.g.Sponheimer & Lee-Thorp 1999) and into the Miocene.
Typically, grasses originally native to hot, arid environments follow the C4 (Hatch-Slack) photosynthetic pathway, and will have d13C values averaging about -12.5‰; trees, shrubs, and grasses from temperate regions,which follow the C3 (Calvin-Benson) photosynthetic pathway, will have d13C values averaging about -26.5‰. In some forested areas, a canopy effect occurs due to incomplete atmospheric mixing, resulting in even more negative carbon isotope ratios (van der Merwe & Medina 1991). Empirical data for large mammals combined with experimental data for rats and mice indicate that bone collagen is enriched about +5‰ relative to diet, although this value is affected by the proportion of protein in the total diet and differences in d13C values between protein and energy sources (van der Merwe 1989; Ambrose & Norr 1993; Tieszen & Fagre 1993). Experimental data on rats also convincingly demonstrates that bone apatite is enriched about +9.5‰ relative to the whole diet, regardless of the mixture or isotopic composition of the foods consumed (Ambrose & Norr 1993), although empirical data for larger herbivores suggest the diet-apatite spacing is about +12‰. Stable carbon isotope analysis is particularly useful in New World dietary studies since maize is often the only C4 plant contributing significantly to human diets; its contribution to bone collagen and to bone apatite may be estimated by interpolation. Some caution is warranted, however, if succulent plants were present, since they utilize the alternative CAM (crassulacean acid metabolism) photosynthetic pathway which results in carbon isotope ratios similar to those of C4 plants. Nevertheless, CAM plants are unlikely to have been major sources of dietary protein, whether consumed directly or indirectly through herbivorous faunal intermediaries.
The carbon isotope ratios of marine and freshwater organisms are more variable, depending on local ecological circumstances, and often overlap with those of terrestrial plants and their consumers. These foods typically have much higher nitrogen isotope values, however, and their high protein content will contribute much more carbon to bone collagen than will maize (about 10% protein) or other plant foods. The analysis of bone apatite,which is derived from all food groups, should allow the identification of just a few percent maize or other C4 resources in an otherwise C3-based diet.
The nitrogen isotope ratios for plants depend primarily on how they obtain their nitrogen - by symbiotic bacterial fixation or directly from soil nitrates - and these values are similarly passed along through the food chain accompanied by an approximately 2-3‰ positive shift for each trophic level, including between mother and nursing infant. Human consumers of terrestrial plants and animals typically have d15N values inbone collagenof about 6-10‰ whereas consumers of freshwater or marinefish, seals and sea lions may have d15N values of 15-20‰ (Schoeninger and DeNiro 1983). While most plants follow either the C3 or C4 photosynthetic pathway and have similar carbon isotope ratios in most ecological settings, nitrogen isotope ratios vary according to rainfall, altitude and other factors (Ambrose1991), and both carbon and nitrogen isotope ratios vary considerably among marine organisms (Schoeninger and DeNiro 1984).
It is critical, therefore, to establish a site-relevant isotopic baseline for interpreting human skeletal data. Analyses of faunal remains provide a good estimate both of the animals themselves and the plants they consume. Establishing an isotopic baseline is particularly important in coastal areas where marine and riverine resources, as well as C4 and/or CAM plants may have been available for direct or indirect consumption by humans.Atmospheric carbon isotope ratios have become depleted by about 1.5‰ since the industrial revolution, so values obtained on modern terrestrial plants and animals must be adjusted accordingly for most archaeological studies.
Other surveys and summaries of stable isotope
analysis
in archaeology may be found in Schwarcz & Schoeninger (1991); van
der
Merwe (1992); Ambrose (1993); Schoeninger 1995; Katzenberg &
Harrison
(1997); Larsen (1997); Tykot
(2004), and Tykot (2006).
My experience in bone chemistry dates to 1990 when I became the manager of the Archaeometry Laboratories in the Anthropology Department at Harvard University. Professor Nikolaas J. van der Merwe, the new director of the laboratory, was the first archaeologist to use stable isotopes, and the methods I learned were adapted from those in use in his laboratory in Cape Town, where he has recently returned as Professor of Natural History. In my lab at USF, we extract bone collagen using well-established laboratory procedures (see also Ambrose 1990). Whole bone is demineralized in 2% hydrochloric acid (72 hrs), base-soluble contaminants are removed using 0.1 M sodium hydroxide (24 hrs each before and after demineralization),and residual lipids are dissolved in a 2:1:0.8 mixture of methanol, chloroform,and water (24 hrs). Freeze-dried collagen pseudomorphs are analyzed for d13C and d15N in continuous flow mode using a CHN analyzer coupled with a Finnigan MAT stable isotope ratio mass spectrometer located at USF. Collagen yields and C:N ratios are used to confirm the integrity of collagen samples.
bone collagen pseudomorphs
the archaeological science laboratory in SCA
Bone apatite and tooth enamel samples are prepared using procedures designed to remove non-biogenic carbon without altering the biogenic carbon isotope values (cf. Koch et al. 1997). Powder samples are obtained by drilling from the center of carefully cleaned bone samples,or from tooth enamel after the surface layer has been removed. Approximately 10 mg of powder are immersed in 2% sodium hypochlorite to dissolve organic components (24hrs for enamel, 72 hrs for bone apatite). Non-biogenic carbonates are then removed in 1.0 M buffered acetic acid for 24 hours. The integrity of apatite and enamel samples is assessed through yields obtained in each stage of the pretreatment process. Samples are analyzed on a second Finnigan MAT mass spectrometer equipped with a Kiel III individual acid bath carbonate system.
The analytical precision for stable isotope ratio
mass
spectrometry is ± 0.1‰ for d13C and ± 0.2‰ for d15N.
I have been involved in a number of collaborative research projects around the world using stable isotope or elemental analysis to assess ancient diets. Preliminary results have been presented at various conferences and some have been published or are in press (see my list of 'publications' on this website). A brief outline of these projects and the results obtained is given below.
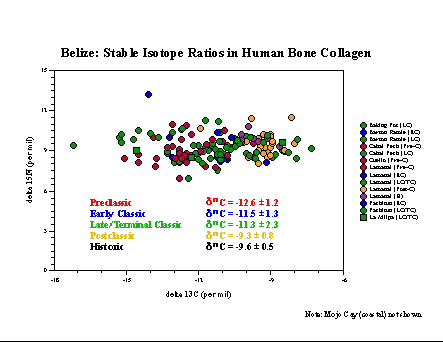
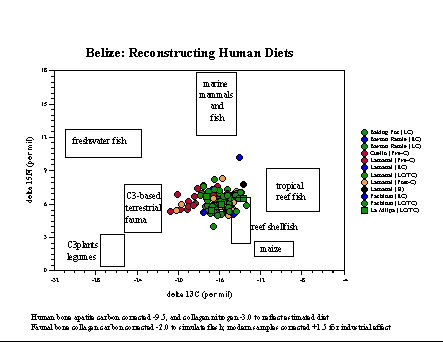
Belize. Skeletal remains from the
Preclassicsite
of Cuello, excavated by Norman Hammond of Boston University, have been
analyzed for bone collagen, bone apatite, and tooth enamel (Tykot
et al.
1996), as have samples from Classic sites at La Milpa (Hammond
excavations)
and the Programme for Belize (Fred Valdez/Richard Adams excavations).
In
conjunction with data published by others, a chronological trend
towards
increasing maize reliance is evident both in bone collagen and bone
apatite.
At Cuello, and probably other sites, some of the maize signature likely
came from the consumption of maize-fed dogs. At La Milpa, diets also
seem
to vary based on social status and residence location.
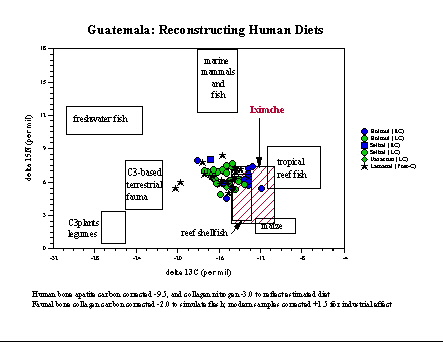
Guatemala. At Iximché in highland
Guatemala,
a number of individuals dating to the Late Postclassic period have been
tested for bone collagen (by David Reed and Stephen Whittington) and
tooth
enamel. The results reveal an extremely high dependence on maize, more
so than most other Guatemalan populations at least of the Late Classic
period, and certainly more so than contemporary populations at Lamanai
in Belize. This high dependence may be related to a combination of
local
ecological as well as socioeconomic factors. Within the group of
individuals
tested, most of whom were decapitated and may have formed a skull rack
(tzompantli), several individuals standout as having
significantly
different diets. We suggest that they may have been warrior-elites from
other communities who consumed different diets either because of
differences
in local ecology, and/or had access to different foods because of their
high status. A full report of our study (
Analysis of Kaqchikel Skeletons: Iximché, Guatemala, S.L.
Whittington
and R. Tykot 2000), which was funded by the Foundation for the
Advancement
of Mesoamerican Studies, Inc., is available on the web.
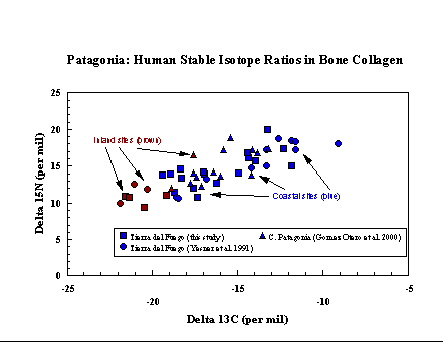
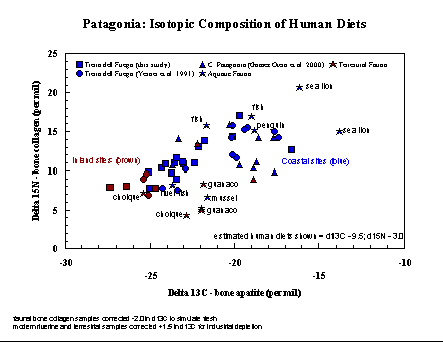
Patagonia (Argentina and Chile). In
southernmost
South America, ethnographic observations of historic groups of
hunter-gatherers
have revealed several discrete subsistence adaptations, including great
dependence on marine foods for those living on the coasts and inlets of
Tierra del Fuego.Growing archaeological evidence, however, has
suggested
at least that this historic pattern does not extend into antiquity.
Isotopic
analysis of bone collagen, bone apatite, and tooth enamel from a large
number of sites in central and southern Patagonia, provided through
collaborations
with Julieta Gomez Otero, Juan Belardi, Ricardo Guichon and Luis
Borrero,
indicates that the coastal populations had a broad range of diets, in
many
cases depending more on terrestrial than marine foods. Some coastal
groups
clearly exploited high-trophic level marine resources such as sea lions
and penguins, while others had more mixed diets which included fish,
mollusks,
guanaco, and land birds. The result appears to be that a continuum of
subsistence
combinations were employed, rather than the discrete selected diets
proposed
by ethnographic observations.
See Gomez
Otero et al. 2000; Borrero
et al. 2001.
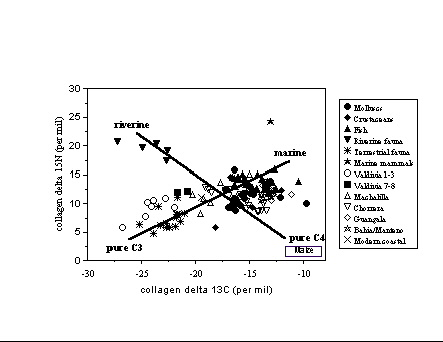
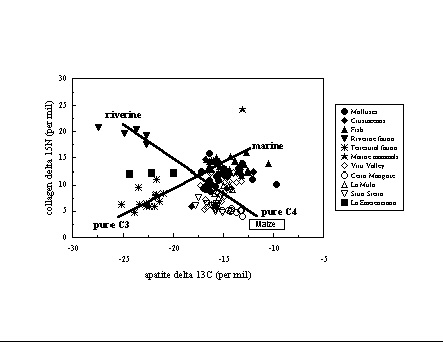
Coastal Ecuador and Peru . The diets
associated
with complex societies on the coasts of Ecuador and Peru have been the
subject of great debate because of the conceived importance of
agriculture-
especially maize - as the basis for sedentary societies and the
emergence
of political hierarchies, and the reported presence of maize pollen and
phytoliths in the region by 5000 BC. The precocious Valdivia culture on
the coast of Ecuador has ceramics and ceremonial sites by 3000 BC,
while
monumental platform structures appear on the coasts of Peru in the 3rd
millennium BC and ceramics shortly afterwards. Analyses of a time
series
of individuals from coastal Ecuador (vander Merwe et al. 1993; Tykot
&
Staller 2002, supplement),
however, suggest that maize is not noticeable in
diets until
the end of the Valdivia period (phases 7-8, about 2000 BC), long after
the appearance of settled villages with ceramics and ceremonial
architecture,and
only became a dietary staple in the second half of the 1st millennium
BC.
In Peru, also, analyses of remains from the Lurin (Mina Perdida,
Cardal,
Tablada de Lurin: Tykot
et al. 2006) and Viru (Ericson et
al. 1989) valleys suggests that maize was not a dietary staple until
well
into the Initial Period. For interpreting coastal diets, where a
combination
of riverine, marine, terrestrial C3 and C4 resources could have been
consumed,
it is important to obtain stable isotope data from both human bone
collagen
and apatite, as well as the available fauna and flora.

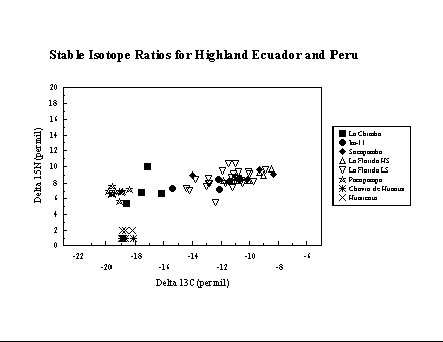
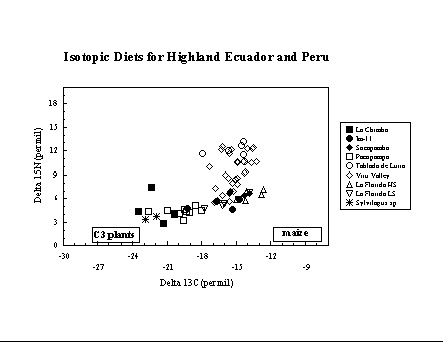
Highland Ecuador and Peru. In highland
Ecuador
and Peru, collaborations with Richard Burger and J. Stephen Athens have
provided the opportunity to examine the introduction and increasing
importance
of maize in the highland Andes. New collagen carbon and nitrogen
isotope
data from Pacopampa and La Chimba support earlier results based on
collagen
carbon from Huaricoto and Chavin de Huantar (Burger and van der Merwe
1990)
which suggest modest importance of maize during the first millennium BC
in the highland Andes. The apatite data, however, suggest that while
maize
was a similar proportion of the whole diet in both regions during the
1st
millennium BC, it was a greater contributor to dietary protein in
highland
Ecuador. In later periods maize became a dietary staple, with
differential
access (e.g. the consumption of maize beer) according to social status
at sites such as La Florida (Ubelaker et al. 1995). Although there
appears
to be an increase in d15N values over time, which could be interpreted
as due to greater consumption of animals (higher trophic level than
plants)
or the availability of small quantities of fish by trade with people
living
at lower elevations, this is most likely an effect of the differences
in
altitude of the sites in our study.
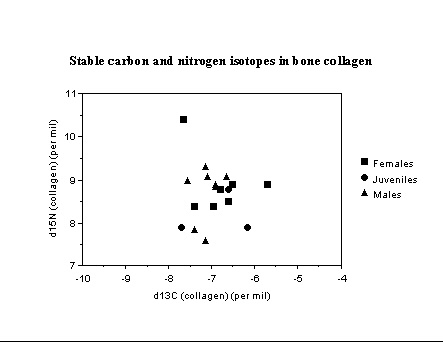
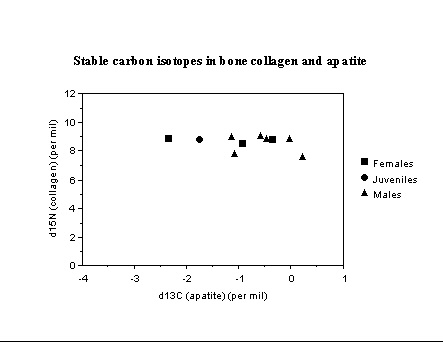
New Mexico. Excavations in the La Plata valley by Debra Martin and colleagues uncovered a number of Anasazi burials (1000-1200 AD), some which received different burial treatments. Stable isotope analysis of bone collagen and bone apatite samples indicate all were heavily dependent on maize. Individuals in flexed, twisted, or 'hurled' burial contexts exhibit gender-based diet differentiation. These results are significant for current discussions of cultural practices and violence/warfare in this region. Two groups of males are suggested by the isotope data, and at least one female appears to have been a recent immigrant to the La Plata valley.
Microsampling of Teeth. Like hair, teeth are incremental growth structures, which presents both advantages and limitations for isotopic analysis. In most cases, archaeologists attempt to be minimally destructive in their analyses, so the removal of a small sample of tooth enamel is required (with the tooth mostly preserved). The diet revealed by isotopic analysis of such a small sample, however, will be very short term, perhaps as little as a few weeks. A series of analyses of sequentially formed enamel layers may then be used to examine short term or seasonal variations in diet, and even to correlate dietary changes with pathologies such as enamel hypoplasia. Preliminary results from our project in collaboration with Douglas Ubelaker shows potentially large isotopic differences within a single human tooth (4 per mil). On the other hand, 'routine' studies in which comparisons are made of individuals (or of multiple teeth from a single individual) from which a single small enamel sample has been tested must allow for the possibility of seasonal dietary variation obscuring variability which otherwise would have been based on eruption age and weaning practices, migration, etc.
 microsampling molar with .75 mm bit
microsampling molar with .75 mm bit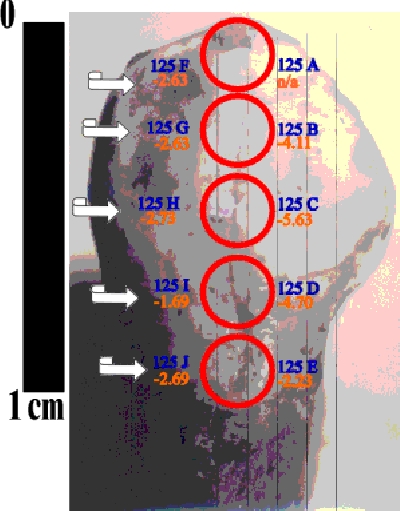 ten d13C measurements on single incisor
ten d13C measurements on single incisor
Elemental Analyses . The elemental composition of bones is also a function of diet, but often more difficult to interpret in archaeological samples (Burton 1996). Elemental ratios of Sr/Ca are a function of trophic level, but also face difficulties in interpretation (see Burton et al. 1999; Wilson 2000). At USF, elemental analyses are obtained by Inductively Coupled Plasma (ICP) Spectroscopy, ICP Mass Spectrometry, DC Plasma Spectroscopy,or Atomic Absorption Spectroscopy. One current project, in collaboration with Francis Thackeray of the Transvaal Museum, is examining trophic levels in baboons and australopithecines from South Africa.
 Jason Wilson using the DCP
Jason Wilson using the DCP  Johan Schijf & the ICP-MS
Johan Schijf & the ICP-MS
For more recent bone chemistry studies in Florida,
Texas, New York, New Mexico, Mexico, Peru, Bolivia, Argentina,
Chile,
Italy, Sardinia,
Malta, Greece, Sweden,
China, and elsewhere, go to the full list of my conference
presentations and publications (in PDF) at http://shell.cas.usf.edu/~rtykot/Pubs.html
Ambrose, S. H. 1990. Preparation and characterization of bone and tooth collagen for isotopic analysis. Journal of Archaeological Science 17(4):431-451.
Ambrose, S. H. 1991. Effects of diet, climate and physiology on nitrogen isotope abundances in terrestrial foodwebs. Journal of Archaeological Science 18: 293-317.
Ambrose, S. H. 1993. "Isotopic analysis of paleodiets: methodological and interpretive considerations," in Investigations of Ancient Human Tissue: Chemical Analysis in Anthropology. Edited by M. K. Sandford, pp. 59-129. Langhorne, PA: Gordon and Breach Science Publishers.
Ambrose, S. H. and L. Norr. 1993. "Experimental evidence for the relationship of the carbon isotope ratios of whole diet and dietary protein to those of bone collagen and carbonate," in Prehistoric Human Bone:Archaeology at the Molecular Level. Edited by J. B. Lambert and G. Grupe, pp. 1-37. New York: Springer-Verlag.
Burger, R. L. and N. J. van der Merwe. 1990. Maize and the origin of highland Chavín civilization: an isotopic perspective. American Anthropologist 92: 85-95.
Burton, J.H. 1996. Trace elements in bone as paleodietary indicators. In M.V. Orna (ed.), Archaeological Chemistry: Organic, Inorganic, and Biochemical Analysis. ACS Symposium Series 625: 327-333. Washington: American Chemical Society.
Burton, J.H., T.D. Price & W.D. Middleton. 1999. Correlation of bone Ba/Ca and Sr/Ca due to biological purification of calcium. Journal of Archaeological Science 26: 609-616.
Ericson, J. E., M. West, C. H. Sullivan and H.W. Krueger.1989. "The development of maize agriculture in the Viru valley, Peru," in The Chemistry of Prehistoric Human Bone. Edited by T. D. Price, pp. 68-104. Cambridge University Press.
Katzenberg, M.A. & R.G. Harrison. 1997. What's in a bone? Recent advances in archaeological bone chemistry. Journal of Archaeological Research 5(3): 265-293.
Koch, P. L., N. Tuross and M. L. Fogel. 1997. The effects of sample treatment and diagenesis on the isotopic integrity of carbonate in biogenic hyroxylapatite. Journal of Archaeological Science 24:417- 429.
Kosiba, S., R.H. Tykot, & D. Carlsson. 2007. Stable Isotopes as Indicators of Change in the Food
Procurement and Food Preference of Viking Age and Early Christian
Populations on Gotland (Sweden). Journal of Anthropological Archaeology
26: 394-411.
Larsen, C. S. 1997. Bioarchaeology: Interpreting Behavior from the Human Skeleton. Cambridge: Cambridge University Press.
O'Connell, T.C. & R.E.M. Hedges. 1999. Investigations into the effect of diet on modern human hair isotopic values. American Journal of Physical Anthropology 108: 409-425.
Schoeninger, M. J. 1995. Stable isotope studies in human evolution. Evolutionary Anthropology 4(3): 83-98.
Schoeninger, M. J. and DeNiro, M. J. 1983. Stable nitrogen isotope ratios of bone collagen reflect marine and terrestrial components of prehistoric human diet. Science 220:1381-1383.
Schoeninger, M. J. and DeNiro, M. J. 1984. Nitrogen and carbon isotopic composition of bone collagen from marine and terrestrial animals. Geochimica et Cosmochimica Acta 48: 625-639.
Schwarcz, H. P. and M. J. Schoeninger. 1991. Stable isotope analysis in human nutritional ecology. Yearbook of Physical Anthropology 34: 283-321.
Sealy, J., R. Armstrong & C. Schrire. 1995. Beyond lifetime averages: tracing life histories through isotopic analysis of different calcified tissues from archaeological human skeletons. Antiquity 69: 290-300.
Sponheimer, M & J.A. Lee-Thorp. 1999. Isotopic Evidence for the Diet of an Early Hominid, Australopithecus africanus. Science 283(5400): 368.
Tieszen, L. L. and T. Fagre. 1993. Effect of diet
quality
and composition on the isotopic composition of respiratory CO2, bone
collagen,
bioapatite, and soft tissues. In Prehistoric Human Bone: Archaeology at
the Molecular Level. Edited by J. B. Lambert and G. Grupe, pp. 121-155.
NewYork: Springer-Verlag.
Tykot, R.H. 2004. Stable Isotopes and Diet: You Are
What You Eat. In M. Martini, M. Milazzo & M. Piacentini (eds.),
Physics Methods in Archaeometry. Proceedings of the International
School of Physics “Enrico Fermi” Course CLIV, 433-444. Bologna, Italy:
Società Italiana di Fisica.
Tykot, R.H. 2006. Isotope Analyses and the Histories
of Maize. In J.E. Staller, R.H. Tykot & B.F. Benz (eds.), Histories
of Maize: Multidisciplinary Approaches to the Prehistory, Linguistics,
Biogeography, Domestication, and Evolution of Maize, 131-142. Academic
Press (Elsevier).
Tykot, R.H. & J.E. Staller. 2002. On the earliest introduction of maize agriculture in coastal Ecuador: new data from La Emerenciana. Current Anthropology 43(4): 666-677.
Tykot, R. H., N. J. van der Merwe and N. Hammond. 1996. "Stable isotope analysis of bone collagen and apatite in the reconstruction of human diet: a case study from Cuello, Belize," in Archaeological Chemistry.Organic, Inorganic and Biochemical Analysis. Edited by M.V. Orna, pp. 355-365. ACS Symposium Series 625, Washington, DC: American Chemical Society.
Ubelaker, D. H., M. A. Katzenberg and L. G. Doyon.1995. Status and diet in precontact highland Ecuador. American Journal of Physical Anthropology 97: 403-411.
van der Merwe, N. J. 1992. Light stable isotopes and the reconstruction of prehistoric diets. Proceedings of the British Academy 77: 247-264.
van der Merwe, N.J. 1989. Natural variation in C13 concentration and its effect on environmental reconstruction using C12/C13 ratios in animal bones. In T.D. Price (ed.), The Chemistry of Prehistoric Human Bone, 105-125. Cambridge University Press.
van der Merwe, N. J., J. A. Lee-Thorp and J. S.Raymond. 1993. "Light, stable isotopes and the subsistence base of Formative cultures at Valdivia, Ecuador," in Prehistoric Human Bone: Archaeology at the Molecular Level. Edited by J.B. Lambert and G. Grupe, pp. 63-97. Berlin: Springer-Verlag.
Whittington, S.L., R.H. Tykot & D.M. Reed. nd. Diet, demography, and death at Iximché, Guatemala: stable isotope analyses of Late Postclassic human skeletal remains from the Kaqchikel Maya capital. Ancient Mesoamerica. Submitted.
Wilson, J.W. 2000. Strontium-Calcium Ratio Analysis and Dietary Adaptations of Early Hominids and Hominoids from South Africa. Senior Honors Thesis, Department of Anthropology, University of South Florida.
Wright, L.E. & H.P. Schwarcz. 1998. Stable
carbon
and oxygen isotopes in human tooth enamel: identifying breastfeeding
and
weaning in prehistory. American Journal of Physical Anthropology 106:
1-18.
Archaeological Science Laboratory Staff and Students
Robert H. Tykot, Director[Back to Homepage] [Back to Research]
Send questions or comments to: R. Tykot